Termorregulação nas aves domésticas.
Naiara Viegas Portella Lemos, Catharine Mendes Costa, Carolina Tonini Prado dos Santos e Ismar Araujo de Moraes.
2021.
Introdução.
As aves são animais homeotérmicos com capacidade de produzir e dissipar calor para manter uma temperatura corporal relativamente constante. No entanto, essa eficiência restringe-se a determinados limites, pois as aves de modo geral não conseguem se ajustar bem a extremos.
A temperatura corporal interna mostra mais variabilidade do que os mamíferos e no frango adulto pode variar 40,6° a 41,7 °C , sendo menor no recém-nascido onde a média é de cerca de 39,7 °C. Os limites máximos de temperatura corporal para a sobrevivência das aves de produção está entre 22,5 ºC e 45-47 ºC, sendo mais tolerantes ao frio do que ao calor.
A literatura em geral demonstra raças menores de galinhas têm uma temperatura corporal mais alta do que raças maiores, que frangos machos têm uma temperatura corporal ligeiramente mais alta do que as fêmeas, provavelmente o resultado de uma taxa metabólica mais alta e de uma massa muscular maior. E ainda que a atividade aumenta a temperatura corporal. Por exemplo, a temperatura corporal das galinhas no chão é mais alta do que a das galinhas mantidas em gaiolas.
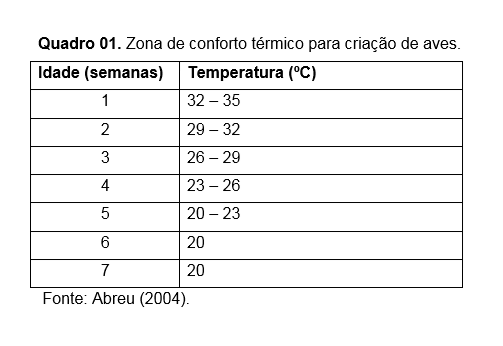
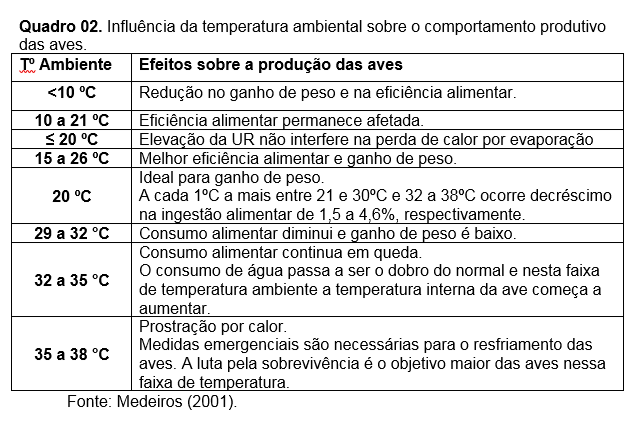
Nos processos de produção que envolve as aves é importante proporcionar facilidade para a termorregulação das aves. É preciso que elas sejam mantidas dentro de sua zona de conforto ou de termoneutralidade e com isso garantir boa produtividade sem a necessidade regulação ativa da temperatura corporal e consumo de energia corporal, além de manter um consumo normal da ração oferecida. Veja os Quadro 01 e 02.
A termorregulação.
Na hierarquia dos sistemas regulatórios, o sistema termorregulador está em um nível superior, e está localizado no sistema nervoso central. O objetivo da regulação da temperatura em animais homeotérmicos na fase de desenvolvimento pós-natal é a manutenção de uma temperatura corporal central estável na maioria das condições. Para realizar isso, o sistema termorregulador emprega todos os sistemas do corpo e integra suas atividades em reações apropriadas e coordenadas. O desenvolvimento pré-natal de mecanismos termorreguladores em animais precoces é benéfico para a rápida maturação da regulação da temperatura na fase inicial pós-eclosão, o que é importante para o desempenho de todo o organismo. (TZSCHENTKE, 2007).
As aves possuem habilidades de regulação da temperatura muito bem desenvolvidas, como capacidade para termogênese regulatória, capacidade de dissipar quantidades consideráveis de calor pela evaporação e mecanismos comportamentais que afetam a transferência de calor entre esses animais e o ambiente. (DAWSON e WHITTOW, 2014). As trocas térmicas do corpo da ave com o meio são realizadas por quatro componentes principais: radiação, condução, convecção e pela evaporação. Esta última constitui um dos mais importantes mecanismos e ocorre na forma da ofegação (polipneia) sem ocorrência de perda evaporativa por sudorese em face da ausência de glândulas sudoríparas. (MELO et al., 2016).
As taxas metabólicas das aves variam ao longo do dia, e com isso a temperatura corporal também passa por alterações, sendo mais alta durante as fases ativas do ciclo diário, tanto em espécies diurnas quanto noturnas. A amplitude térmica tende a variar inversamente com o tamanho do corpo do animal (ASCHOFF e POHL, 1970 apud DAWSON e WHITTOW, 2014).
As alterações fisiológicas que acontecem no organismo na busca pelo equilíbrio térmico são possíveis devido a percepção da temperatura do ambiente, que ocorre graças aos termorreceptores. Esses sensores de temperatura localizam-se em diferentes partes do corpo e transmitem as informações para o cérebro, que então dá início aos mecanismos termorregulatórios (KLEIN, 2014).
Segundo Necker (1972, 1973 apud DAWSON e WHITTOW, 2014) e Gentle (1987, 1989 apud DAWSON e WHITTOW, 2014), há mais evidências de fibras aferentes para o frio e poucas demonstrações de receptores para quente. Entretanto, a atividade dos receptores para o quente é alta e há um aumento da atividade estática na faixa de 25 a 45°C. A atividade dos termorreceptores frios nas aves é menor que em receptores frios de mamíferos.
Assim como todos os demais animais homeotérmicos, as aves produzem calor metabólico de modo contínuo haja vista os processos naturais tais como alimentação, movimentação dentro e fora de galpões, digestão, reprodução e outros. Para manter a temperatura estável e dentro da faixa de termotolerância as aves podem lançar mão de mecanismos fisiológicos ou comportamentais voltados para a geração de calor (termogênese) ou perda de calor (termólise).
Mecanismos termogênicos e manutenção do calor
Nas extremidades nuas das aves, o fluxo sanguíneo para os tecidos pode ser mantido sem que haja perda de calor devido a estruturas vasculares especiais que permitem a ocorrência de transferência de calor do sangue arterial que se direciona a uma extremidade para o sangue venoso que retorna ao coração. Essas estruturas consistem em veias que se agrupam em torno de uma artéria, e permitem que o calor do sangue arterial, que seria perdido na superfície do corpo, se mantenha no organismo (DAWSON e WHITTOW, 2014). Esse tipo de troca de calor entre o sangue quente arterial que desce e sangue frio venoso que retorna às partes internas corpo do animal é denominado de mecanismo de troca de calor por contracorrente.
Do inglês, as “brood patches” correspondem a áreas de pele nua e sem penas na parte inferior do peito das aves e podem ser consideradas como uma característica importante na termorregulação, pois são áreas muito vascularizadas. Essas áreas são também importantes para garantir a transferência de calor corporal para os ovos ou filhotes, favorecendo a manutenção da espécie (DAWSON e WHITTOW, 2014). Proteger as “brood patches” de substratos frios será uma forma de manter o calor corporal e expô-la a estes pode garantir a perda do calor quando necessário.
O comportamento também desempenha um importante papel nas relações térmicas desses animais, permitindo-lhes economizar energia, conservar água e reduzir o estresse térmico. Normalmente ele envolve alguma mudança de posição e postura ou até mesmo permanecer repousado, quiescente. Por exemplo, durante períodos frios e com muito vento, as aves podem procurar abrigos ou evitar agitar suas penas, que em contato com o vento gelado, reduzindo a temperatura corporal. Ainda, algumas espécies de coruja se encurvam, reduzindo a área de superfície e consequentemente a perda de calor. E quando no frio, os pássaros dobram sua cabeça sob as penas escapulares para conservar sua temperatura. (DAWSON e WHITTOW, 2014). De modo interessante, a adoção de uma postura de sono, na qual as aves mantêm-se encolhidas e acocoradas, reduz a perda de calor por meio de cristas, barbelas e narinas (BROOM e FRASER, 2010).
Mecanismos temolíticos
O calor transportado para a superfície da pele pode ser perdido por meios não evaporativos, que pode ocorrer por condução, convecção e irradiação (WHITTOW, 1986 apud DAWSON e WHITTOW, 2014). A perda de calor convectiva é um componente significativo na equação do equilíbrio térmico para as aves por conta de seu movimento pelo ar. Entretanto, é a radiação que mais contribui para a perda de calor não evaporativa (DAWSON e WHITTOW, 2014).
O calor que esses animais podem perder em ambientes quentes através da evaporação total é de grande importância, podendo ser igual ou exceder o valor produzido no metabolismo em muitas espécies. Como a maior parte do corpo da ave é coberta por penas, o calor e a água devem atravessar a plumagem antes de serem perdidas para o ambiente, tornando a evaporação pela superfície da pele mais difícil (DAWSON e WHITTOW, 2014). As penas são um ótimo isolamento em climas frios, mas inibem a perda de calor em climas quentes.
Foi observado que a principal forma de perda de calor evaporativo ocorre nas regiões nasal e superior da traqueia através da respiração, que é muito utilizada por aves em circunstâncias desafiadoras e extremas (LASIEW-SKI, 1972 apud DAWSON e WHITTOW, 2014). Durante o estresse térmico nas aves, a frequência respiratória eleva de forma acentuada, podendo chegar a um aumento de seis a sete vezes ciclos respiratórios por minuto (LUDDERS, 2017). Esse aumento envolve respiração de boca aberta e denomina-se polipneia térmica, sendo responsável por aumentar significativamente a evaporação respiratória (DAWSON e WHITTOW, 2014).
O saco gular é uma bolsa de pele elástica localizada na região cervical de algumas espécies de aves, abaixo do bico. Alguns animais, como pelicanos, abutres, codorna, pombos e corujas (DAWSON e WHITTOW, 2014), complementam a perda de calor a partir da vibração rápida e intensa dessas áreas, o que promove aumento do fluxo de ar no espaço morto respiratório. (KLEIN, 2014; EWART, 2021). Essa vibração é produzida pela contração e relaxamento de forma alternada do aparelho hióide e, em pombos, esse mecanismo é favorecido pela inflação rítmica ou pulsação do esôfago (GAUNT, 1980; BAUMEL et al., 1983 apud DAWSON e WHITTOW, 2014).
A literatura aponta que esse mecanismo parece ter vantagens em comparação com a polipneia térmica. Isso porque o custo energético para vibrar o saco gular é consideravelmente menor do que para mover a estrutura toracoabdominal, resultando em maior eficiência de resfriamento (DAWSON e WHITTOW, 2014). Além disso, o sistema de controle respiratório atua de forma a maximizar a ventilação respiratória das vias respiratórias superiores, aumentando a perda evaporativa de água e resfriando o corpo, porém sem hiperventilar os brônquios, garantindo a ausência de alteração na difusão dos gases e do pH do sangue arterial (LUDDERS, 2017).
Mudanças comportamentais também podem auxiliar na perda de calor para o ambiente, como procurar ambientes que protejam da radiação solar direta (BROOM e FRASER, 2010). Especialmente em pássaros pequenos, a cabeça (principalmente próximo aos olhos) tende a ser uma região de perda substancial de calor (VEGHTE E HERREID, 1965; HILL et al., 1980 apud DAWSON e WHITTOW, 2014). Ainda, aumentar a superfície corporal ao manter as asas afastadas do corpo quando a temperatura encontra-se elevada também é uma forma de reduzir a temperatura corporal (NASCIMENTO e SILVA, 2009; BROOM e FRASER, 2010).
Referências bibliográficas
ABREU, P. G.; ABREU, V. M. N. Conforto Térmico para Aves. Comunicado Técnico EMBRAPA. Ministério da Agricultura Pecuária e Abastecimento. Concórdia, 2004.
BROOM, D. M.; FRASER, A.F. Comportamento e Bem-Estar de Animais Domésticos. 4ed. Editora Manole, 2010. 452p.
DAWSON, W. R.; WHITTOW, G. C. Regulation of Body Temperature. In: SCANES, C. G. Sturkie’s Avian Fisiology. 6ed. Academic Press. 2014. 685p. cap 14, p. 344-379.
EWART, S.L. Termorregulação. In: KLEIN, B. Cunningham Tratado de Fisiologia Veterinária. 6 ed. GEN Guanabara Koogan, 2021.
KLEIN, B. Termorregulação. In: ____. Cunningham Tratado de Fisiologia Veterinária. 5 ed. Rio de Janeiro: Elsevier, cap. 53, p. 1419 -1447. 2014. 1599 p.
LUDDERS, J.W. Respiração nas Aves. In: REECE, W. Dukes Fisiologia dos Animais Domésticos. 13 ed. GEN Guanabara Koogan, 2017.
MEDEIROS, C. M. Ajuste de modelos e determinação de índice térmico ambiental de produtividade para frangos de corte. 2001. 115 p. Tese (Doutorado em Construções Rurais e Ambiência). – Universidade Federal de Viçosa. Viçosa, MG.
MELO, A. S et al. Relação temperatura e nutrição sobre o desempenho de galinhas poedeiras. Pubvet, v. 10, p. 795-872, 2016.
NASCIMENTO, S. T.; SILVA, I.J O. As perdas de calor das aves: entendendo as trocas de calor com o meio. Revista AviSite. 2009
TZSCHENTKE, B. Atingimento de termorregulação conforme afetado por fatores ambientais. Ciência Avícola, v. 86, p. 1025-1036, Ed. 5, 2007.