A AUDIÇÃO DOS ANIMAIS
Referência: LIMA, A.A.; MORAES, I. A. A audição dos animais. Webvideoquest de Fisiologia Veterinária. 2017.
INTRODUÇÃO
A audição é um importante órgão do sentido dos animais que lhe permite maiores chances de sobrevivência seja na natureza ou nos ambientes já modificados pelo homem. Ela é importante para detectar diferentes perigos, entre eles os predadores e os veículos motorizados, para responder às verbalizações de seus semelhantes em comportamentos de acasalamento além de uma variedade de outras necessidades da vida (Strain e Myers, 2006).
ANATOMIA
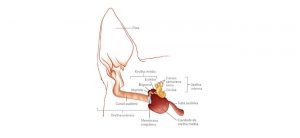
Fonte: Princípios de fisiologia animal, 2010.
As partes anatômicas envolvidas com a audição são a orelha externa, orelha média e orelha interna (Dyce et. al, 2010). A maioria dos vertebrados não mamíferos não possui uma orelha externa óbvia, e os peixes não apresentam as orelhas externa e média; porém, todos os vertebrados possuem uma orelha interna. (Moyes, 2010).
Orelha externa
Dela fazem parte a aurícula e o meato acústico externo (canal auditivo). A aurícula tem seu formato determinado pela cartilagem auricular e movimentos determinados pelos músculos auriculares que pode volta-la em direção à origem do som. As aurículas direita e esquerda podem mover-se de forma independente, de modo que cada uma possa se concentrar em sons isolados. O meato acústico externo, por sua vez, é o canal que comunica a base da aurícula com o tímpano. Ele possui partes cartilaginosas e ósseas, e revestimento interno rico em glândulas sebáceas e ceruminosas tubulares (Dyce et al, 2010).
Orelha média
Está inserida no osso temporal, e nela encontram-se um pequeno espaço cheio de ar conhecido como cavidade timpânica, além da membrana timpânica e de uma cadeia ossicular formada pelo martelo, bigorna e estribo (Dyce et. al, 2010). Segundo (Balbani e Montovani, 2008) essa cadeia ossicular é exclusividade dos mamíferos, sendo encontrado nas aves apenas um único ossículo chamado de columela.
A cavidade timpânica da orelha média é revestida por uma mucosa delgada que se comunica com a nasofaringe por meio da tuba auditiva responsável pela equalização das pressões dos dois lados das membranas do tímpano. As tubas se abrem temporariamente no bocejo ou na deglutição. Em equinos, a parede membranosa da tuba auditiva evagina-se por meio de uma falha ventral no suporte cartilaginoso, formando a ampla bolsa gutural (divertículo da tuba auditiva). Na parede lateral da cavidade timpânica encontra-se a membrana timpânica e na parede medial encontram-se as duas janelas (do vestíbulo e da cóclea), fechadas no seu estado natural, através das quais os estímulos mecânicos produzidos por ondas sonoras entram na orelha interna, para tradução em impulsos nervosos. A janela do vestíbulo une a cavidade timpânica ao vestíbulo da orelha interna. A janela da cóclea conduz a cavidade da cóclea (Dyce et al., 2010).
Orelha interna
É completamente envolvida pela parte petrosa do osso e consiste em um sistema fechado de minúsculos ductos e cavidades membranosas conhecidas por sua complexidade como labirinto membranoso (Dyce et al., 2010).
Do ponto de vista fisiológico, o ouvido interno é dividido em uma parte acústica — a cóclea, e uma parte não acústica — o órgão vestibular (Santos, 1975). A cóclea se apresenta na forma de ductos paralelos e espiralados repletos de líquido e está dividida em três compartimentos pela membrana basilar e membrana de Reissner, os quais são denominados rampa vestibular, rampa média e rampa do tímpano. A rampa vestibular e do tímpano são repletas de perilinfa e a rampa média pela endolinfa ( Strain e Myers, 2006 ). Segundo Guyton e Hall (2017) a endolinfa contém alta concentração de potássio e baixa de sódio, e a perilinfa tem sua composição quase idêntica à do líquido cefalorraquidiano. Segundo os autores existe um potencial elétrico entre a endolinfa e a perilinfa, chamado de potencial endococlear, que é gerado por secreções contínua de íons potássio positivos para a rampa média. A importância do potencial endococlear é que os topos das células ciliadas se projetam, através da lâmina reticular, sendo banhados pela endolinfa da rampa média, enquanto a perilinfa banha os corpos das células ciliadas (Guyton e Hall, 2017).
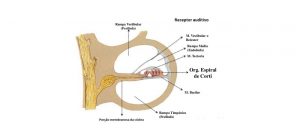
A membrana de Reissner é tão fina e tão móvel que não obstrui a passagem de vibrações sonoras da rampa vestibular para a rampa média. A importância da membrana de Reissner é de manter um tipo especial de líquido na rampa média, que é necessário para a função normal das células ciliadas (Guyton e Hall, 2017).
A rampa média tem como componente de tecido conjuntivo a lâmina basiliar que é importante na percepção do som. As células formam o órgão espiral, onde originam os impulsos nervosos produzidos pelos sons recebidos pela orelha externa (Dyce et al., 2010). Estendendo-se pelo ducto, encontra-se o órgão de Corti que contém células epiteliais sensoriais possuidoras de cílios imóveis ( Strain e Myers, 2006 ).
O órgão de Corti é então o responsável pela percepção do sentido da audição. Ele está situado sobre a membrana basilar na rampa média e contém receptores sensoriais: uma fileira de células pilosas internas e três ou quatro de células pilosas externas. Cada célula pilosa tem múltiplos estereocílios, dispostos nas células pilosas externas em um zigue-zague que aponta para fora do modíolo, o eixo em torno do qual a cóclea forma as espirais ( Strain e Myers, 2006 ). De acordo com Guyton e Hall (2017), as fibras nervosas, estimuladas pelas células ciliadas, levam ao gânglio espiral de Corti, que se situa no modíolo (centro) da cóclea. As células neuronais do gânglio espiral enviam axônios para o nervo coclear e, depois, para o sistema nervoso central no nível da parte superior do bulbo.
Apenas os mamíferos possuem cóclea verdadeira. As aves e crocodilianos apresentam um ducto coclear com estrutura aproximadamente reta que também possui o órgão de Corti e membrana basilar (Balbani e Montovani, 2008).

A TRANSMISSÃO E PERCEPÇÃO DO SOM
Os estímulos mecânicos produzidos pelo som são transformados em impulsos nervosos na orelha interna através do órgão de Corti (Santos, 1975; Dyce et. al, 2010) .
A transmissão de ondas sonoras pela cavidade timpânica é mediada pelos três ossículos auditivos que fazem vibrar a membrana timpânica ligada ao manúbrio do martelo. O manúbrio age com ação de alavanca e coloca em movimento a base do estribo, fazendo vibrar o líquido da orelha interna e consequentemente as células neurorreceptoras no ducto membranoso da cóclea, e assim o som é percebido ( Dyce et. al, 2010).
As ondas trafegam rapidamente ao longo da parte inicial da membrana basilar, mas ficam progressivamente mais lentas quando se afastam em direção à cóclea. A causa dessa diferença é o alto coeficiente de elasticidade das fibras basilares, perto da janela oval e do coeficiente progressivamente menor ao longo da membrana. Essa transmissão inicial rápida das ondas permite que os sons com alta frequência cheguem longe o suficiente, na cóclea, para se propagarem e se separarem na membrana basilar. Sem essa rápida transmissão inicial, todas as ondas de alta frequência se agrupariam, mais ou menos, no primeiro milímetro da membrana basilar, e suas frequências não poderiam ser discriminadas (Guyton e Hall, 2017).
Um tom puro que entra na cóclea desencadeia uma onda de pressão constante que produz uma deflexão máxima na membrana basilar em um ponto ao longo do seu comprimento, variando a localização sistematicamente de acordo com a frequência do tom. A consequência é que frequências diferentes são detectadas de maneira excelente em diferentes pontos ao longo da cóclea, sendo as altas frequências (ondas curtas) mais bem detectadas na base perto da janela oval e as baixas frequências (ondas longas) no ápice da cóclea. O resultado desse código de frequência é que as células pilosas ao longo da membrana basilar estão arranjadas de acordo com a sensibilidade à frequência (organização tonotópica), que possibilita a discriminação da tonalidade. Os sons complexos consistem em misturas de sons de várias frequências de modo que a resposta coclear a eles consiste nas respostas a vários componentes de tons puros do complexo sonoro, sendo todos transmitidos para o sistema nervoso central (Strain e Myers, 2006 ).
Espécie
|
Nº de voltas cocleares
|
Variação audível da frequência (Hz)
|
Cão
|
3,25
|
67-45.000; 200-36.000
|
Gato
|
3,0
|
45-64.000; 125-60.000
|
Bovino
|
3,5
|
23-35.000; 140-22.000
|
Equino
|
2,5
|
55-33.500; 200-22.000
|
Ovino
|
2,25
|
100-30.00; 400-37.00
|
Morcego
|
2,25
|
2.000-110.00
|
Galinha
|
–
|
125-2.000
|
Fonte: Strain e Myers, 2006
Intensidade do som
A informação acerca da intensidade do som é extraída na cóclea por vários mecanismos. A membrana basilar não apenas vibra seletivamente em um local correspondente a frequência do som, como também o faz em uma amplitute proporcional à intensidade do som. A maior distorção de determinada célula pilosa resulta em maior magnitude do potencial receptor, o qual resulta em maior frequência de potenciais de ação produzidos nas células ganglionares espirais pelas células pilosas. O limiar para dor em resposta a sons muito altos parecem variar de acordo com a espécie e é provável que também sejam dependentes da frequência ( Strain e Myers, 2006 ).
Vias neurais da audição
A informação coclear sobre o som é levada para o tronco cerebral nos processos dos neurônios bipolares do gânglio espiral, os quais se tornam a parte auditiva do VIII nervo craniano. As fibras passam do córtex auditivo primário para o secundário (ou de associação). A organização tonotópica de frequências ao longo da membrana basilar da cóclea é mantida em todas as estruturas auditivas até o córtex cerebral. No córtex auditivo primário do lobo temporal, as baixas frequências são mapeadas em áreas rostrais, e as altas frequências, em áreas caudais. O córtex auditivo primário funciona extraindo aspectos acerca do som — frequência, intensidade e padrão —, e o córtex auditivo secundário processa a informação, em vez de reconhecer os sons (Strain e Myers, 2006).
Keifer (1999) relata que em mamíferos as fibras nervosas do gânglio espiral de Corti vão aos núcleos cocleares (dorsal e ventral) na região superior do bulbo, onde todas as fibras fazem sinapse com neurônios de segunda ordem. Daí as fibras se projetam através do corpo trapezoide para o lado oposto do tronco, onde seguem ao núcleo olivar superior medial e lateral; as fibras que não cruzam dirigem-se ao núcleo olivar do mesmo lado. Seguindo pelo lemnisco lateral a maioria das fibras terminam no núcleo do lemnisco lateral e parte delas passam diretamente indo ao colículo inferior (mesencéfalo). O colículo inferior é o ponto de junção sináptica para praticamente todas as fibras auditivas. Do colículo inferior a via se dirige ao núcleo ou tálamo, onde ocorre nova retransmissão sináptica de todas as fibras, seguindo para o córtex auditivo, localizado em sua maior parte no giro superior do lobo temporal. A informação proveniente dos sinais sonoros começa a ser dissecada em níveis cerebrais baixos (núcleos cocleares).
Percepção da localização do som
Os neurônios do tronco cerebral nos núcleos olivares, bem como os neurônios em estruturas mais altas, possibilitam a localização de uma fonte sonora, ao comparar as diferenças entre os ouvidos na intensidade do som e na latência de chegada. As diferenças na latência de chegada são melhores para localizar os sons de baixa frequência. As diferenças na intensidade são melhores para localizar os sons de alta frequência, porque a cabeça os absorve mais que os de baixa frequência, enfatizando qualquer diferença na intensidade de alta frequência entre as orelhas ( Strain e Myers, 2006 ).
De acordo com Guyton e Hall (2017), os mecanismos neuronais para detectar a direção do som acontece da seguinte maneira: O núcleo olivar superior se divide em duas partes, o núcleo olivar superior medial e o núcleo olivar superior lateral. O núcleo lateral está relacionado com a detecção da direção da qual o som está vindo, presumivelmente, por simples comparação da diferença de intensidades do som que chega aos dois ouvidos e pelo envio de sinal apropriado ao córtex auditivo, para estimativa da direção. O núcleo olivar superior medial, contudo, tem mecanismo específico para detectar o intervalo de tempo entre os sinais acústicos que entram nos dois ouvidos. Esse núcleo contém grande número de neurônios que têm dois dendritos principais, um se projetando para a direita e outro para a esquerda. O sinal acústico do ouvido direito invade o dendrito direito, e o sinal do ouvido esquerdo invade o dendrito esquerdo. A intensidade de excitação de cada neurônio é muito sensível ao intervalo de tempo específico entre os dois sinais acústicos dos dois ouvidos. Os neurônios próximos a borda do núcleo respondem, de modo máximo, a intervalo de tempo curto, enquanto os próximos da borda oposta respondem a intervalo de tempo longo; os intermediários respondem a intervalos de tempo intermediários.
AUDIÇÃO NAS AVES
Balbani e Montovani (2008) relatam que na maioria das aves a abertura do meato acústico externo tem poucos milímetros de diâmetro e é recoberta por penas. A membrana timpânica é grande em relação ao tamanho da cabeça na maioria das espécies, especialmente nas corujas. As tubas auditivas se abrem em um óstio faríngeo único, situado na linha média.
A cóclea das aves tem cerca de 1/10 do comprimento da cóclea dos mamíferos, mas possui maior densidade de células ciliadas e boa capacidade de regeneração após trauma sonoro ou exposição a ototóxicos.
Leia mais: Audição nas aves: https://repositorio.ufsc.br/bitstream/handle/123456789/80952/139919.pdf?sequence=1
AUDIÇÃO NOS MORCEGOS
Kandel et al. (2014) mostra que, conforme estudado por Nobuo Suga e colaboradores, muitos morcegos emitem pulsos eco localizadores com dois componentes. Um componente de frequência constante (FC) inicial consiste em vários sons harmonicamente relacionados. Esses harmônicos são emitidos de forma estável por dezenas a centenas de milissegundos. O componente de FC é seguido por um som cuja frequência decai abruptamente, denominado componente de frequência modulada (FM). Os sons de FM são usados para determinar a distância até o alvo. O morcego mede o intervalo entre o som emitido e o eco que retorna, o qual corresponde a uma determinada distância, com base na velocidade relativamente constante do som.
AUDIÇÃO NOS INVERTEBRADOS
Segundo Strain e Myers (2006) a audição é limitada aos vertebrados e insetos. Balbani e Montovani (2008) informaram que o órgão auditivo dos insetos tem morfologia variada e pode localizar-se em diferentes partes do corpo do animal, como nos segmentos abdominais (nos gafanhotos), nas patas anteriores (nos grilos e esperanças), nas antenas (em moscas) ou nas asas (em mariposas). O órgão timpânico geralmente se desenvolve a partir de dilatações da traquéia e da parede do corpo do inseto. As vibrações do tímpano se transmitem a terminações nervosas das cavidades aéreas adjacentes. A maioria dos insetos possui pequeno número de células sensoriais auditivas quando comparada com os animais superiores. Por essa razão sua habilidade se limita à localização da fonte sonora e à detecção de uma estreita faixa de frequências. As exceções são as cigarras Tettigetta josei e os gafanhotos Bullacris membracioides que possuem até 2.000 receptores sensoriais em cada órgão auditivo, além de inúmeros interneurônios.
REFERÊNCIA BIBLIOGRÁFICA
DYCE, K. M.; SACK, M. O., WENSING, C.J. G. Os órgãos dos sentidos. In: Tratado de anatomia veterinária . 4ª Edição. Rio de Janeiro: Elsevier, 2010. Cap. 9, p. 346-350.
STRAIN, G.M.; MYERS, L.J. Audição e equilíbrio. In: REECE, W.O. Dukes Fisiologia dos Animais Domésticos . 12ª ed. Rio de Janeiro, 2006. Editora: Guanabara Koogan, cap.47, p. 793-804.
MOYES, C.D., SCHULTE, P. M. Princípios de Fisiologia Animal . 2ª Edição. Ed. Artimed, 2010.
SANTOS, J. A. Patologia especial do ouvido. In: Patologia especial dos animais domésticos – mamíferos e aves. 1ª Edição. Ed. Interamericana, 1975.
KANDEL, E. R., SCHWARTZ, J. H., JESSELL, T.M., SIEGETHAUM, S. A., HUDSPETH, A. J. Princípios de Neurociências . 5ª Edição. Ed. Artmed, 2014.
KEIFER, C.L. Sensibilidade auditiva de neuronios mesencefalicos em pombos (Columba livia). 1999. Texto disponível em: <https://repositorio.ufsc.br/bitstream/handle/123456789/80952/139919.pdf?sequence=1>. Acesso em: 18 jul. 2017.
BALBANI , A. P. S. ; MONTOVANI , J. C. . Aspectos filogenéticos da audição. Revista Brasileira de Medicina ORL. Controvérsias & Interfaces , v. 3, p. 37-45, 2008.

